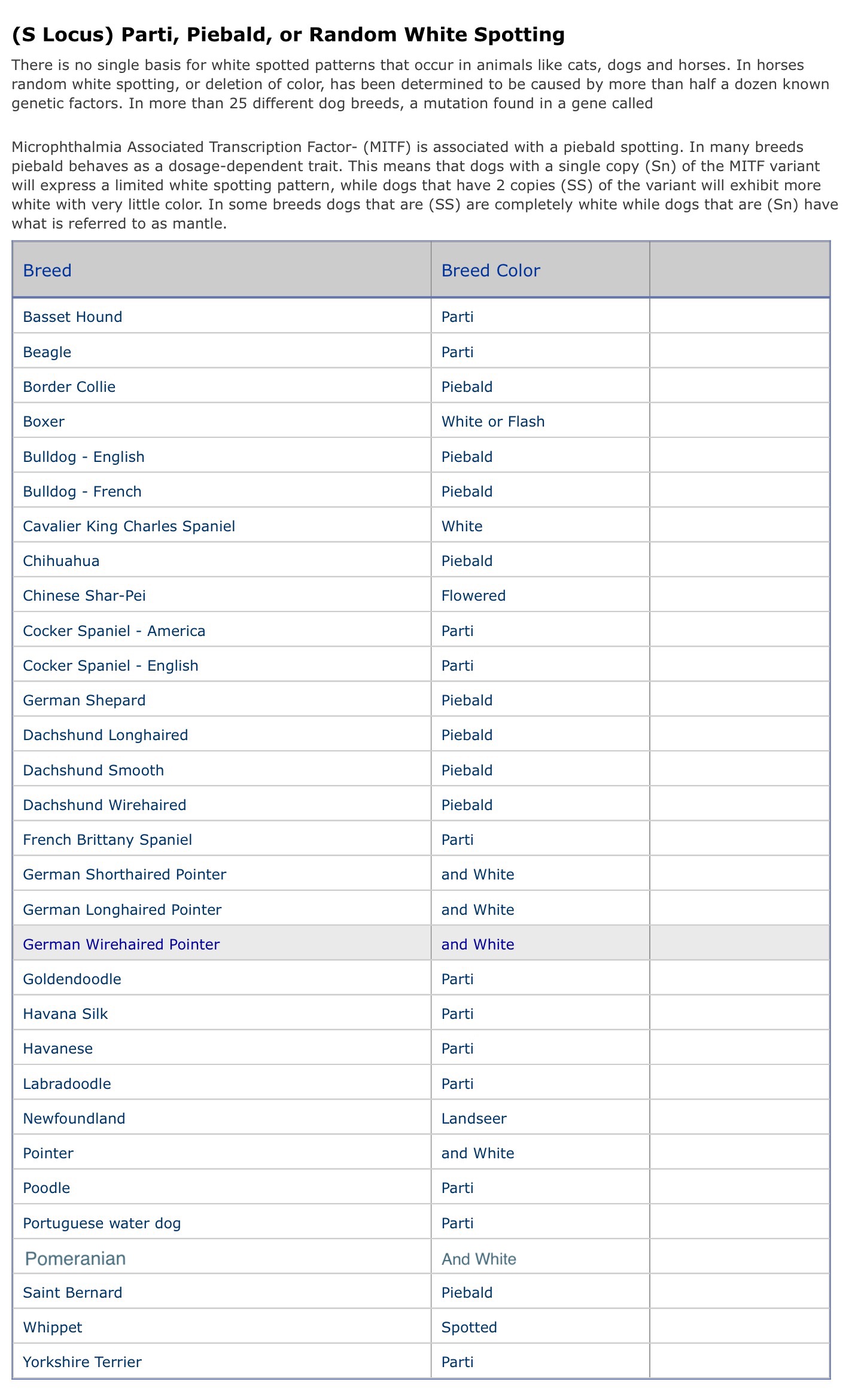
There are a number of intermediate genes where the mutation apparently produces a partly active form of tyrosinase. Some C alleles known in other mammals are:
- C full color, allows full expression of whatever pigment is prescribed by other genes. Most dogs are CC.
- c, chinchilla or silver, when present in double dose removes most or all of the phaeomelanin pigment with only a slight effect on black pigment. This is named after a small fur-bearing South American rodent called the chinchilla. Black and silver replacing black and tan, or a wolf-like color without the extra banding (see aw, above) may also be due to a cchcch genotype. Dogs with very light tan probably are cchcch or something similar. Liver dogs show lightening even of eumelanin pigment, and the "deadgrass" color of the Chesapeake Bay Retriever is thought to be due to a bbcchcchgenetic makeup. The possibility of other, rufous modifiers affecting the shade of phaeomelanin pigment needs to be kept in mind, as does the possibility of more than one form of chinchilla in the dog - rabbits are thought to have three.
- ce, extreme dilution, has also been proposed for the dog. This gene may be part of the makeup of some "white" dog breeds where the white color is due to extreme dilution of tan. The West Highland White Terrier may be ceceee. A cross to a black and tan breed would be interesting from the point of view of color genetics. Eyes may be lightened in some species, but this is doubtful in dogs.
- ch, Himalyan, is not known to occur in the dog. In homozygous form, it makes the formation of eumelanin dependant on the temperature of the skin. Thus a genetically solid black animal will have reduced black on the extremities (seal brown) and an almost white color on the body. The effect on tan/orange pigment is confusing - the tan in agouti hairs is removed, but that resulting from the orange gene in cats (not in dogs) remains intense on the extremities. There is reason to suspect that this gene, as well as some forms of chinchilla, also affects the organization of the brain, particularly in the neural pathways from the eyes to the brain. There may be a reason for Siamese cats to be cross-eyed. Eyes are normally blue or pink.
- cp, platinum, is optically similar to Ice White but retains very slight tysonase activity and in the mouse is described as retaining some luster in the coat as opposed to the pure white seen in Ice White .
- c, albino, is known to occur in the dog RARELY , though possible candidates for mutations to c have been recorded. As mentioned above, the c gene cannot produce working tyrosinase, and a cc individual cannot produce melanin pigment.
As seen from the above, C is known to have a number of different forms and effects. The usual assumption is that dogs have at least one mutant allele, cchwhich when homozygous lightens phaeomelanin (yellow) pigment to cream and more weakly affects liver and black. A second proposed allele, ce may be responsible for further reduction of cream to white in some breeds, or modifying alleles may be responsible for the further lightening in these cases. While some forms of C modify eye pigment. C appears to be fully dominant over any of the other alleles, the dominance relationship between the others generally goes in the direction of more color incompletely dominant over less color, the heterozygote generally resembling but not necessarily identical to the homozygote with more pigment
D, the dilution series. This, again, is a relatively simple series, containing D (dominant, full pigmentation) and d (recessive, dilute pigment). In contrast to C, which has its strongest effect on phaeomelanin, or B, which effects only eumelanin, D affects both eumelanin and phaeomelanin pigment. It is thought to act by causing the clumping of pigment granules in the hair. Like B, it often affects skin and eye color.... blue" is a term often used to describe dd blacks. If a solid liver dog also is dd, the result is the silvery color known as Lavendar .
While dd acting on black or liver is a part of the genotype of several breeds, dd acting on sable is relatively rare. For one thing, the action of dd on phaeomelanin has been described as a flattening or dulling of colorCanine Color Genetics
BY
Canine Coat Color
VetGen DNA CHROMAGENE Coat Color Testing
For many years geneticists and breeders have been aware of several locations on the chromosomes, or loci, which are responsible for the color patterns we see in dogs and other mammals. As with all genetic traits, every animal inherits one copy of each locus from each of its parents. Each of these loci is responsible for one or more traits either independently, or in conjunction with another locus. All of them act on the pathways that produce the two major pigments, phaeomelanin and eumelanin, or affect the distribution of those pigments.
The combined effect of all these loci (Agouti, Brown, Extension, etc.) is the color of the dog. Due to the complex interactions of these genes, it is possible for dogs to carry hidden colors which may appear in their offspring. Over the past several years, scientists at VetGen and elsewhere have determined the actual genes associated with many of these loci, and identified the mutations responsible for the different versions (alleles) of these genes. The fruit of this work are the tests available for many of the common coat colors and traits.
The diagram below illustrates the relationships among the major loci (A, B, E and K) involved in determining coat color. The sections that follow describe the role these loci and others play in the coat color tests that VetGen offers. These tests demonstrate that while a dog may exhibit certain color or colors, it may also be carrying other hidden colors in its genetic code that can show up in later generations.
How do the A, B, E, and K loci affect each other in determining coat color?
The accompanying graphic helps to illustrate the interactions of the genes at these four loci in a hierarchy in terms of their role in coat color. If a circle is filled with color, it means the color of the dog has been determined at that point. If a circle is still white, it means information about an additional gene is required.
The first locus to look at is the E locus. The gene at this locus is responsible for black masks when present as well as most shades of yellow and red. Any dog that is "ee" will be some shade of yellow to red, and everything happening at the A, B, and K loci will be hidden until the next generation. If the dog has any E or Em alleles, then it will not be yellow and we must look next at the K locus.
There are three versions, or alleles, of the K locus: KB, kbr, and ky. If a dog has even a single copy of KB (KBKB, KBky, KBkbr) it will be solid colored in the pigmented areas, and we go directly to the B locus to determine color. Everything happening at the A locus in these dogs is hidden until the next generation. If a dog is kyky, it will not be brindled, and we go next to the A locus to see which alleles are expressed. If a dog is kbrkbr, or kbrky, it will nearly always be brindled and we look next at the A locus to see the background color and pattern of that brindling.
The A locus has at least four alleles. There are direct tests available for Ay (fawn or sable),"a" (recessive black), and "at" (tan points). There is no direct test for the Wild type (think wolf pattern) allele which is designated a .Any dog which has at least one copy of Ay (and no KB) will be fawn or sable, either with or without brindling.Any dog that is "aa" (and no KB) will be black. Any dog that is atat or ata (and no B) will have tan points, either with or without brindling.
The next stop is the B locus. Any dog which is "bb" will be have brown fur in those areas that would otherwise be black. This holds true for both solid colored and agouti-patterned animals.
The D locus (not shown in the diagram) can alter the intensity of pigment. Animals which are "dd" exhibit grey or blue fur in place of black, and light tan or "Isabella" in place of brown. This tan is similar to some AY shades but lacking any banding or black tips on individual hairs.
AVAILABLE TESTS
E locus
The E locus is responsible for the black mask seen in many breeds, and more significantly, for the presence of the yellow to red coats of many dogs. The gene involved is known as MC1-R, which has at least three versions affecting the appearance of the dog, E, Em, and e. Dogs with two copies of e will be yellow, orange or red in their pigmented coat regardless of their genotype at all the other loci
Research at VetGen and independently at the University of Saskatchewan has identified two new alleles in the E locus, Eg and Eh. These mutations are responsible for a reverse mask or widow's peak appearance in the "domino" Afghan Hound and "grizzle" Saluki (Eg), as well as the "sable" English Cocker Spaniel (Eh). Research continues to determine if they are responsible for similar appearance in additional breeds.
Test for "e"
Analysis proves absence or presence of the mutation typically responsible for yellow, lemon, red, cream, apricot and some white in at least the following breeds and all dogs with these breeds in their lineage:
Afghan, Australian Cattle Dog, Australian Shepherd, Border Collie, Beagle, Brittany Spaniel, Cardigan Welsh Corgi, Chinese Shar-pei, Chow Chow, Cocker Spaniel, Dachshund, Dalmatian, Doberman Pinscher, English Cocker Spaniel, English Setter, English Springer Spaniel, Field Spaniel, Flat-Coated Retriever, Foxhound, French Bulldog, German Longhaired Pointer, German Shepherd, German Shorthaired Pointer, German Wirehaired Pointer, Irish Setter, Labrador Retriever, Lowchen, Pointer, Pomeranian, Poodle, Pudelpointer.
Test for Em
Analysis reveals whether a dog with a mask has one or two copies of this version of the extension locus. Animals with a single copy can produce offspring with or without a mask, while those with two copies will only produce masked offspring. The test may also be applied to black dogs where it may not be possible to tell if there is a mask. It may be present in the following breeds and all dogs with them in their lineage:
Afghan, Belgian Shepherd, Boxer, Bull Mastiff, Cairn Terrier, Cardigan Welsh Corgi, Chihuahua, Chinese Shar-pei, English Bulldog,German Shepherd, Great Dane, Greyhound, Lakenois, Pekinese, Pug, Saint Bernard, Saluki, Staffordshire Bull Terrier,Whippet.
Test for Eg and/or Eh
Analysis reveals the absence or presence of the mutations responsible for "grizzle" in Salukis and "domino" in Afghans (Eg) or "sable" and "dirty red" in English Cocker Spaniels (Eh).
B Locus
The B locus is responsible for the presence of brown, chocolate, or liver animals. It is also responsible for nose color. The gene associated with this locus is known as TYRP1. In breeds where the A locus does not come into play, any animal that has at least one B allele (and is not "ee"), will be black in pigmented coat. Those dogs, which have two copies of any of several b alleles will be brown. There are at least three such b alleles. Regardless of other loci, any animal with at least one B allele will have a black nose and pads, while those with any two b alleles will have a liver nose and pads.
Test for b
This test analyzes whether an animal has 0, 1 or 2 copies of the mutations typically responsible for brown, which is also known in some breeds as liver, chocolate, sedge, and less frequently, red. There are three primary "b" mutations that are responsible for nearly every liver or chocolate dog. A notable exception is the French Bulldog where in addition to these three mutations, there is a fourth cause of chocolate that has yet to be identified.
It can be present at least, but not exclusively, in the following breeds:
Australian Cattle Dog, Australian Shepherd,Bedlington Terrier, Border Collie, Brittany Spaniel,Cardigan Welsh Corgi, Chihuahua, Chinese,Shar-pei, Cocker Spaniel, Curly Coated Retriever,Dachshund, Dalmatian, Doberman Pinscher,English Cocker Spaniel, English Setter,English Springer Spaniel,Field Spaniel, Flat-Coated Retriever, Fox Terrier, French Bulldog,German Longhaired Pointer, German Shorthaired Pointer, German Wirehaired Pointer,Labrador Retriever, Lowchen,Miniature Pinscher,Newfoundland Pointer, Pomeranian, Poodle, Portuguese Water Dog, Pudelpointer,Scottish Terrier, Skye Terrier,Weimaraner, Wirehaired Pointing Griffon.
Also:
Any dogs that contain these breeds in their lineage.
K Locus
The K locus plays a pivotal role in coat color. This locus is a relative newcomer in our understanding of canine color, and includes traits formerly attributed by some to other genes.
The dominant allele in the series is KB, which is responsible for self-coloring, or solid colored fur in pigmented areas. This trait was formerly attributed to the Agouti (A) locus as AS, but recent breeding studies had shown this not to be the case.
There are two other alleles, kbr, and ky. KB is dominant to both kbr and ky, while kbr is dominant only to ky. kbr is responsible for the brindle trait and for a long time had been considered to belong in the E locus. Recent breeding studies had also shown this to be incorrect. The recessive allele, ky, allows the basic patterns of the A locus to be expressed. So too does the kbr allele, but with brindling of any tan, fawn, or tawny areas.
Any animal with at least one KB allele will be self-colored.
Any animal with at least one kbr allele, and no KB allele will be brindled on agouti background (see A locus).
Any animal with two ky alleles will show agouti patterns (see A locus).
The mutations responsible for these alleles were identified and described primarily by Sophie Candille in the laboratory of Dr. Greg Barsh at Stanford University.
Test for KB and ky
Vetgen can presently test for these two alleles. In some breeds, where no brindle is present, this represents a complete analysis of the locus. An example would be the Pug. In breeds where the breed standard disqualifies all but self-colored dogs, testing for these two alleles is once again all that is needed. Any animal with two KB alleles cannot produce anything except self-colored offspring. A prime example here is the Labrador retriever. In breeds where many variations are allowed, these tests can help predict the probability of potential litters to include fawn, sable, tawny, tan point, tricolor or recessive black puppies.
| KB KB | self-colored (solid color in pigmented areas) |
| KB kbr | self-colored (solid color in pigmented areas) |
| KB ky | self-colored (solid color in pigmented areas) |
| kbr kbr | allows A locus to express (tan point, tricolor, fawn, sable, tawny) with brindling |
| kbr ky | allows A locus to express (tan point, tricolor, fawn, sable, tawny) with brindling |
| ky ky | allows expression of agouti patterns without brindling |
A Locus
The A locus is responsible for a number of common coat patterns in the dog. Expression of all of them requires any combination of two ky or Kbr alleles at the K locus, and at least one E or Em allele at the E locus. The gene involved is the Agouti gene, and variations in it are responsible for fawn and sable dogs (Ay), wild type (Aw), tan points (at), and recessive black(a).
Test for Ay
Analysis proves absence or presence of the mutation typically responsible for fawn or sable. In fawn/ sable dogs this test shows if other agouti alleles are present but hidden (only one copy of Ay). It also demonstrates how many copies of this allele are hidden in dogs, which cannot express agouti types (KBKB, KBkbr, KBky, at the k locus and/or "ee" at the E locus).
Test for “a”
Analysis shows whether a black dog is black due to “recessive black,” or the more common black at the K locus. It also reveals whether a non-black animal carries “recessive black.” Examples of breeds:
German Shepherd Dog, Shetland Sheepdog, Belgians (Tervuren, Malinois,Lakenois, Groenendael).
Test for “at”
Vetgen is now offering a test for a mutation that is found in all tan point, phantom, tricolor, and "black/tan, chocolate/tan and liver/tan" dogs. This mutation, at, was identified and reported by researchers at the University of Saskatchewan.
In order to produce “at” pups, the pups need to inherit both an at and a ky or kbr allele from both parents, but no KB . In many breeds where the occasional tan point dog is viewed as unwanted, the k test is still warranted since the majority of dogs have two copies of this at mutation, but do not express it because of the presence of an overriding KB.
The “at” mutation is also found in recessive black dogs, but does not cause recessive black (a). In breeds that do not have recessive black (most breeds), this test alone will indicate the presence of “at”. In breeds where recessive black is present, “at” can be determined by using this test in conjunction with the recessive black test, or by our previous process of elimination approach of testing for Ay and recessive black.
It should also be noted that recessive black is present at a very low frequency in more breeds than we once thought (ie French Bulldogs, Poodles and Tibetan Mastiff).
Testing for this mutation along with Ay and recessive black (a) also allows for the identification of Aw alleles in those breeds where it is present.In breeds where only the Ay and at alleles are present, the A,sup>y test can be used to see if the fawn/sable dog is Ay/Ay (homozygous) or only has one Ay (heterozygous). If it only has one, the other allele must be at (ie Afghans,Collies,Cardigan Welsh Corgi, Dachshund, Norwich Terrier, Staffordshire Terrier).
In breeds where only Ay, a and at alleles are present, both the Ay test and the "a" test need to be performed. Any alleles unaccounted for by these two tests will be at. For example, if a dog is Ay/Ay both alleles are accounted for. If a fawn/sable dog only has a single Ay, then the other allele must either be an "a" or an at and this can be determined by running the recessive black ("a") test or the at test (ie Shetland Sheepdog, Belgian Shepherd (Tervuren, Malinois, Lakenois, Groenendael).
D Locus
The D locus is the primary locus associated with diluted pigment, which results in coats that would otherwise be black or brown instead showing up as gray, or blue in the case of black, and pale brown or Isabella in the case of brown. The melanophilin gene has recently been shown to be responsible, but not all of the dilute causing mutations have been identified yet.
M Locus
The M locus is responsible for the merle and double merle color patterns seen in some breeds. The mutation which causes merle in all of its forms has been identified. VetGen cannot offer a test for merle because it is patented elsewhere.
Coat Length
While it is not a color trait, the length of a dog's coat is of interest to many. It has recently been demonstrated that in many breeds, the FGF5 gene is responsible for whether a dog has a long coat (rough or fluffy), or a short (smooth) coat. The test Vetgen offers detects the presence or absence of the recessive allele that results in long coats when present in two copies, and as such allows dogs with short coats that carry a hidden "long coat" allele to be detected. In addition to the original coat length mutation, research at VetGen has identified some new mutations present in northern breeds (Akita, Chinook, Siberian Husky) that are responsible for the "woolly" long coat. These new tests are offered exclusively through VetGen.
Furnishings
Furnishings refer to the longer facial hair around the eyebrows, moustache, and beard commonly seen in many breeds, including the wirehaired breeds. Presence of furnishings is dominant to the unfurnished version of the gene, which depending on breed may also be referred to as satin, or sleek. VetGen offers a test to see if a furnished dog carries the recessive unfurnished trait, which is considered unfavorable in some breeds.
Curly
The gene responsible for curly coat has also been identified. Most breeds are fixed for either curly or not-curly, but in breeds where there is variation, dogs may now be tested to see if they carry zero, one or two copies of the curly or non-curly versions of the KRT71 gene.
Canine Color Testing- B Locus
|
|||||||
|
Description Because TYRP1 is only associated with eumelanin, this mutation only has an effect on dogs that are "EE" or "Ee" at the E-locus. Dogs that are "ee" only produce phaeomelanin in their coats, so a mutation at the B-locus will not have an effect on their coat color. However, eumelanin is still produced in the foot pads and noses of "ee" dogs, so the B-locus still has an effect on these areas. Dogs that are "eebb" will have a brown nose and foot pads, rather than black.
Cost
B-Allele Results:
|
 TYRP1,
or tyrosinase-related protein 1, is a protein that plays a role
in the synthesis of the pigment eumelanin. In the dominant form
of this gene, or the "B" allele, normal eumelanin is produced
in the coat, and the dog's coat appears black in color. A mutation
in the TYRP1 gene can occur causing a change in function which dilutes
the black color pigment to a brown color. This mutated gene is known
as the "b" allele. When a dog is homozygous for the mutation,
meaning he has 2 copies of the recessive allele, the dog's coat
color will be brown in color. This color can also be referred to
as liver, chocolate, or in some breeds, red.
TYRP1,
or tyrosinase-related protein 1, is a protein that plays a role
in the synthesis of the pigment eumelanin. In the dominant form
of this gene, or the "B" allele, normal eumelanin is produced
in the coat, and the dog's coat appears black in color. A mutation
in the TYRP1 gene can occur causing a change in function which dilutes
the black color pigment to a brown color. This mutated gene is known
as the "b" allele. When a dog is homozygous for the mutation,
meaning he has 2 copies of the recessive allele, the dog's coat
color will be brown in color. This color can also be referred to
as liver, chocolate, or in some breeds, red.
|
|||||||||||||||||||||||||||||||||||||||||
|
Animal Genetics, Inc. |

 of
the "a" allele. This is also the case for dogs that are bicolor, but
are negative for K Locus. This is generally the cause of a solid black
German Shepherd. The "a" allele is sometimes referred to as the
recessive black gene. Because is the most recessive, for a dog to
express this phenotype, he must have two copies of the "a" allele. A
recessive black dog will always pass on the "a" allele to all offspring.
of
the "a" allele. This is also the case for dogs that are bicolor, but
are negative for K Locus. This is generally the cause of a solid black
German Shepherd. The "a" allele is sometimes referred to as the
recessive black gene. Because is the most recessive, for a dog to
express this phenotype, he must have two copies of the "a" allele. A
recessive black dog will always pass on the "a" allele to all offspring.
|
||||||||||||||||||||||||||||
|
Animal Genetics, Inc. |

|
|||||||||
|
Animal Genetics, Inc. |
 The
MLPH gene codes for a protein called melanophilin, which is responsible
for transporting and fixing melanin-containing cells. A mutation
in this gene leads to improper distribution of these cells, causing
a dilute coat color. This mutation is recessive, so two copies of
the mutated gene, or "d" allele, are needed to produce
the dilute coat color.
The
MLPH gene codes for a protein called melanophilin, which is responsible
for transporting and fixing melanin-containing cells. A mutation
in this gene leads to improper distribution of these cells, causing
a dilute coat color. This mutation is recessive, so two copies of
the mutated gene, or "d" allele, are needed to produce
the dilute coat color.Canine Color Testing- K Locus
|
||||||
|
Description The Dominant Black gene consists of three different alleles, or variants. The first allele, which is dominant, is noted as "KB," or dominant black. The Dominant Black allele is actually a mutation that does not allow the agouti gene to be expressed. Because this mutation is dominant, a dog only needs to have one copy of the mutation to suppress the agouti locus. A dog that has one or two copies of the Dominant Black allele will only express his base coat color, which is determined by the B-Locus and E-Locus. He will not express any colors that occur from the agouti gene, such as "black and tan" or "tricolor." The second allele is known as the "brindling" allele, and is represented as "Kbr." The Kbr allele is a separate mutation that still allows the agouti gene to be expressed, however, causes brindling of the agouti patterns. The agouti gene represents several different colors, such as fawn/sable, tricolor, tan points, or recessive black.The Kbr allele is recessive to the KB allele, however, it is dominant over a third allele, Ky. Thus, for a dog to express the brindle pattern, he must be either Kbr/Kbr or Kbr/Ky. Dogs that are KB/Kbr will not appear brindle, but can still pass on that allele and potentially produce brindle offspring. The third allele is represented as "Ky." This allele allows the agouti gene to be expressed without brindling. When a dog is Ky/Ky at the K-locus, the agouti locus determines the dog's coat color. For example, a dog that is Ay/Ay at the agouti locus could be fawn/sable. If that same dog is KB/KB at the K-locus, the agouti locus will be hidden, and his coloration will be determined at the B- and E- loci. However, if that same dog is Ky/Ky at the K-locus, he will then be able to express agouti, and will be fawn/sable. At this time, there is no direct test for the "Kbr" allele, although it can generally be inferred through testing for the presence of the Dominant Black allele, as well as through phenotypes of the parents and offspring. Testing for the Dominant Black mutation determines if the dog is able to express agouti phenotypes, however, it is limited in that it will not tell you if the dog will be brindled.
Cost
KB-Allele Results:
|

 |
 |


Tests Offered (click color name below for more information):
MC1R (Melanistic Mask & Black/Red) |
Agouti |
Brown |
Dilute (Blue) | Dominant Black(K locus)| Piebald, Parti or Random White Spotting |Coat Length, Curl and Furnishings|
Order these tests on MyVGL
BackgroundMammals have two pigments that are the basis of hair color: eumelanin (black) and pheomelanin (red or yellow). The gene involved in the production of these pigments in many species including dogs is Melanocortin 1 Receptor (MC1R) which is also called Extension. Other genes modify these pigments to produce the variety of colors and patterns found in the domestic dog. The Brown gene, Tyrosinase-Related Protein 1 (TYRP1), is a modifier that dilutes black pigment to brown but does not affect red pigment. Other genes involved in dog coat color include Agouti (ASIP) which organizes the distribution of black and red pigments, Beta-defensin (CBD-103) which is unique to dogs and responsible for dominant black (K locus), and Dilute (MLPH) which dilutes black and red pigments. Other genes that add white patterns and dilute colors are also present in dogs but are specific to certain breeds. Below are expanded descriptions of the genetic tests offered by the Veterinary Genetics Laboratory for dog coat color.
Allow 5-10 business days for test results.

 |
|
Dilution - D gene
homozygous recessive form (dd)
homozygous recessive form (dd)
This recessive gene has a diluting effect on both eumelanin and phaeomelanin. When present in the homozygous recessive form (dd) it dilutes black/brown eumelanin to blue, and red to cream.
If he is black (B/B or B/b) and gets d/d (which is the homozygous recessive form), that will dilute the black color to blue.
|
lightened. If the Merle gene is added, the color will dilute to blue merle. This merle is true blue (d/d) merle and is much lighter than the black merle. In the true blue merle, all of the black patches have been diluted to blue, gray, and pewter colors (no black color is found). The nose, lips and eye rims will be also be gray.
If he is brown/chocolate (b/b) and gets D/D or D/d, his color remains brown/chocolate.
|
If he is brown/chocolate (b/b) and gets d/d, then his brown is diluted to a dull, flat silvery-brown color called dilute brown (called Lilac ). The nose, lips and eye rims are a rosey-gray color.
|
| If two dogs are mated that are both b/b d/d (Lilac), all of the puppies will be Lilac. |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 B Locus (Brown): This gene determines or selects for a black or brown dog. When this gene is in its dominant form (BB or Bb) the dog is black. When this gene is in its homozygous recessive form (bb) it has a lightening effect on the eumelanin only and the dog is brown.
B Locus (Brown): This gene determines or selects for a black or brown dog. When this gene is in its dominant form (BB or Bb) the dog is black. When this gene is in its homozygous recessive form (bb) it has a lightening effect on the eumelanin only and the dog is brown.

FYI:
When brown (b/b) is expressed, it means that the final step in eumelanin production has not been completed and
the pigment remains brown. The brown color is not a genetic defect.
When the alleles are in the homozygous or heterozygous dominant form of B/B or B/b, the color and pigment
(nose, eye rims and lips) remains (or directs the color to be) black.
When the alleles are in the homozygous recessive form (b/b), the color and pigment will be brown. This just means
that the final step in eumelanin production of changing brown to black did not occur. Phaemelanin (yellow/red
[e/e]) is not affected. BUT, in the e/e colored dog, if the dog is also b/b; they will be either red or yellow and will
have brown pigment (nose, eye rims and lips). e The pigment granules produced by "bb" are smaller, rounder in
shape, and appear lighter than pigment granules in "B" dogs. The iris of the eye is also lightened.
DNA specific tests can now be carried out for the presence of most of the colour alleles, particularly where one wants to know if there are unwanted dilution factors hiding within individuals. While it can look very complicated, try to understand the subject and thus produce the colours you want from matings and not waste litters with incorrect colours. Knowledge of your proposed breeding pairs’ colour genetics can help maximise desired colour combinations.
General information
Melanocytes are the cells that produce skin and hair colour and they are derived from neural crest cells. These cells arise along the back very early in foetal development and then give rise to a number of cell types, including a large proportion of the peripheral nervous system. If there is a decrease in the number of neural crest cells, other cell types are favoured, leading to a reduction in melanocyte formation (see below). The melanoblasts (immature colour cells) migrate from the dorsal midline over the surface of the body, so the last areas to be reached are the feet, chest and muzzle (ie where you are more likely to see white toes, etc) As you would see in a "Mis-mark " Pomeranian for example.
Neural crest cells also form part of the nervous system for the inner ear and eye. Animals selected for extreme white spotting (eg. Dalmatians) can have hearing and/or vision problems in other extreme white patterns (merle series). Dalmatian deafness is thought to be as a result of the absence of melanocytes in the stria vascularis of the inner ear.
Melanoctyes produce melanin by the action of alpha MSH. Large amounts of MSH results in Eumelanin (mainly Tyrosine) which produces black or derivatives of black (blue, chocolate, brown, liver). Restricted amounts of MSH result in Phaeomelanin (contains varying amounts of cystine and tyrosine) which produces reddish brown or yellowish tan.
#Control of melanoctye function is intricate and many loci(genes) have mutations which affect components of melanogenic control mechanisms.
There are about 10 recognised "loci" of different colour genes. Each locus can have a variable number of alleles that can influence the colour outcome (dilution, pattern, dominant or recessive effects).
All dogs carry these genes, many of which are in a fixed (homozygous) form eg. Gordon Setters, Elkhounds etc. In these breeds virtually all loci are fixed and there is very little colour variation across the breed. In other breeds some loci are fixed, while others have a degree of variation (number of alleles) present at other loci. Not all breeds carry all the possible alleles at each locus.
Current research - Many of the colour loci are being extensively studied to precisely locate the position of the various genes on the correct chromosome, and further, which alleles actually occur at that site and how they affect the colour outcomes available. From this, DNA markers may be developed, and thus allow DNA colour testing in various breeds prior to mating. The DNA research that is being done at this time is going to continue to change our understanding of colour genetics - its a fairly dynamic field - it is certainly not fixed in concrete.
Gene / allele designation - The way I write the alleles is to put the loci first as a capital. If only 2 alleles occur (dominant and recessive) then eg. D means the dominant allele, d the recessive allele. Where there are more than 2 alleles for a loci, I find it easier to keep the capital as the indicator for the loci, followed by a smaller letter indicating the allele. (If one has a fancy typewriter/computer, one could do the allele in smaller letters above the series loci letter.) If there are several alleles at a loci, these are usually written in decreasing order of dominance (according to current knowledge).DID YOU KNOW......................a Pomeranian puppy can be born almost black and end up a very light creme color three years later as an adult ? It is fascinating to see the changes in Pomeranian coat color as they grow from birth to adulthood . Some great examples are documented superbly by
There are several coat colors that can be associated with health concerns.
http://skyway.usask.ca/~schmutz/conditions.html

One such gene is MLPH or the D locus. Several, but not all dogs with a d/d genotype have Color Dilution Alopecia and/or Black Hair Follicular Dysplasia. Although d/d dogs could be blue, grey, pale brown or a dilute red, it is unlikely that the orange or cream of Pomeranians is due to this genotype since blue or grey are not listed as allowed colors and I have not seen photos of such dogs. In addition, there are black-and-cream Pomeranians which have a decrease in phaeomelanin on their undersides to cream but no decrease on their backs which remain black. It is therefore likely that it is the I gene that causes the variation in shade in orange (e/e) dogs.

Another gene that causes major health concerns is AP3 which causes Grey Collie Syndrome. Luckily this has only been identified in Collies since it is lethal to pups.
Yet another gene in the "blue" family that can cause health issues is merle. Merle can not be seen in dogs with an e/e genotype. This e/e genotype occurs commonly in Pomeranians since orange, red and white probably account for the majority of Poms. The problem is that M/M (homozygous merle) dogs are always deaf based on our studies. We recently genotyped 24 mostly white Australian Shepherds and all tested M/M (based on the Clark et al. 2006 PNAS published test) and all were deaf. A proportion of these dogs were also blind in one or both eyes since microphthalmia is another common side effect in M/M dogs. Although in many breeds it is possible to educate breeders to never breed two merle dogs together this advice is not possible to follow in Pomeranians since e/e dogs would not show the merle pattern. It would therefore be necessary instead to advise all persons who breed a merle dog to use only a black or sable mate or to have DNA testing done on their red, orange or white mate prior to breeding to be sure it did not carry merle.

This is a photo of a homozygous merle puppy with microopthalmia (small eyes). Note how small the eyes are compared to those of a normal puppy. One or both eyes can show reduction in size, and one or both eyes can be missing completely (anopthalmia).
To determine whether or not a Pomeranian carries the merle gene, a DNA test is now available. This test is only available through GenMark at a cost of $95.00 - see http://www.genmarkag.com/home_companion.php. Because the merle gene can be carried by an apparently normal dog, those that want to avoid this cruel gene will now have to add the expense of merle DNA testing to the other health tests that they already perform. Even those that want to produce merle puppies will have to perform this test to ensure that they are not breeding two merle gene carriers to one another since you cannot always tell by a dog's appearance if it has merle in its genetics
Although albinism is rare in dogs, it does occur. The side effects associated with albinism have not been well documented since the phenotype is so rare and may be caused by different genes in different breeds, each with different side effects. This should not be confused with white dogs that have deeply pigmented nose leather and pads however which do not have health concerns. The occasional white dog with pigmented leather is M/M but usually dogs of this genotype has splashes of mottled color on their body.
ALBINO
Sheila Schmutz, Ph.D.
Professor, University of Saskatchewan
Prior to 1996 the AKC standard read: red, orange, cream, sable, white, parti-color, black and black-and-tan. The main advantages of specifying coat colors are:
provide educational information to owners and breeders
provide information to judges
provide information to geneticists who are then able to tailor a DNA test package which identifies the alleles for these specific colors
identify which colors are excluded for either health, aesthetic or historical reasons
I would advocate this approach to most breeds. Pre-1996 the major genes and alleles causing variation in Pomeranian coat color were likely:
A (agouti) = agouti signalling protein (ASIP)
ay fawn (cream to yellow to red with darker tips)
sable (some solid black hairs intermingled amongst reddish hairs)
at black-and-tan or brown-and-tan
E (extension) = melanocortin receptor 1 (MC1R)
EM melanistic mask
E eumelanin can be produced
e only phaeomelanin produced
K (from blacK......"dominant black") (gene identified but not published yet)
K solid black, brown or blue (eumelanin pigmentation only)
k expression of agouti alleles that express phaeomelanin possible
I (dilues only phaeomelanin) = ? gene (Co-Dominant)
I intense, not diluted
i co-dominant decrease in intensity
S (spotting) = ? gene (in pigs this is KIT but may not be just one gene in dogs)
S solid colored
si Irish spotting
sp piebald or random spotting
Sheila Schmutz, Ph.D.
Professor, University of Saskatchewan
DID YOU KNOW THAT YOU CAN DO Color Testing FOR - A Locus ? YOU CAN FIND OUT IF YOU'R POMERANIAN IS REALLY A WOLF SABLE | |||||||||||||
ASIP, or agouti signalling peptide, is a gene affects pigmentation of coat color in dogs. This protein interacts with the melanocytes to stimulate production of melanin. This gene exists in many forms, or alleles, in dogs. The first of these alleles is generally known as AW, or the wild-type allele. This appears as a "wolf-grey" type of pattern. There is no direct test for this allele, and it is thought that this is the most dominant allele. The second allele, known as AY, is a dominant allele responsible for fawn or sable pigment. A dog only needs 1 copy of the AY allele to be either fawn or sable, depending on the breed. This allele is responsible for the coat color seen in dogs such as Pugs or French Bulldogs. In terms of dominance, it is thought that AW is the most dominant, followed by AY. A third allele is the "tan points" allele, or at. There is no direct test available for this allele at this time, however, it is expressed in dogs that do not carry the AY allele. This phenotype is common in dogs such as Doberman Pinschers or Geman Shepherds. The fourth allele is known as "recessive black," or the a-allele. This variant of the agouti gene causes a dog that does not carry the dominant black gene to be solid black. The recessive black allele is responsible for all-black German Shepherds, for example. The recessive black allele is recessive to all other alleles, meaning that the dog must carry two copies of the "a" allele to express this pattern. The Agouti gene is only visible in dogs that do not carry the dominant black gene. The dog can still carry any of the agouti alleles, however, this effect is hidden by the dominant black gene.
Cost
AY-Allele Results:
a-Allele Results:
|
WANT TO KNOW IF YOU REALLY HAVE A BLUE POMERANIAN | |||||||
The MLPH gene codes for a protein called melanophilin, which is responsible for transporting and fixing melanin-containing cells. A mutation in this gene leads to improper distribution of these cells, causing a dilute coat color. This mutation is recessive, so two copies of the mutated gene, or "d" allele, are needed to produce the dilute coat color. This mutation affects both eumelanin and phaeomelanin pigments, so black, brown, and yellow dogs are all affected by the dilution. A dilute black dog is generally known as "blue," though other names do vary for different breeds, such as charcoal or grey.
Cost
D-Allele Results:
|
Coat coloration is controlled by several different genes in dogs. One of these gene is referred to as "Dominant Black." The presence or absence of this gene determines if a dog can express "agouti" color patterns.
The Dominant Black gene consists of three different alleles, or variants. The first allele, which is dominant, is noted as "KB," or dominant black. The Dominant Black allele is actually a mutation that
does not allow the agouti gene to be expressed. Because this mutation is dominant, a dog only needs to have one copy of the mutation to suppress the agouti locus. A dog that has one or two copies of the Dominant Black allele will only express his base coat color, which is determined by the B-Locus and E-Locus. He will not express any colors that occur from the agouti gene, such as "black and tan" or "tricolor."The second allele is known as the "brindling" allele, and is represented as "Kbr." The Kbr allele is a separate mutation that still allows the agouti gene to be expressed, however, causes brindling of the agouti patterns. The agouti gene represents several different colors, such as fawn/sable, tricolor, tan points, or recessive black.The Kbr allele is recessive to the KB allele, however, it is dominant over a third allele, Ky. Thus, for a dog to express the brindle pattern, he must be either Kbr/Kbr or Kbr/Ky. Dogs that are KB/Kbr will not appear brindle, but can still pass on that allele and potentially produce brindle offspring.
The third allele is represented as "Ky." This allele allows the agouti gene to be expressed without brindling. When a dog is Ky/Ky at the K-locus, the agouti locus determines the dog's coat color. For example, a dog that is Ay/Ay at the agouti locus could be fawn/sable. If that same dog is KB/KB at the K-locus, the agouti locus will be hidden, and his coloration will be determined at the B- and E- loci. However, if that same dog is Ky/Ky at the K-locus, he will then be able to express agouti, and will be fawn/sable.
At this time, there is no direct test for the "Kbr" allele, although it can generally be inferred through testing for the presence of the Dominant Black allele, as well as through phenotypes of the parents and offspring. Testing for the Dominant Black mutation determines if the dog is able to express agouti phenotypes, however, it is limited in that it will not tell you if the dog will be brindled.
DO YOU REALLY HAVE A BRINDLE |
Description |
Animal Genetics currently offers a test for the K-Locus to determine how many copies of the dominant "KB" allele a dog carries.
Cost
$40.00 US for the KB-allele test.
Sample Collection
Collect sample using buccal swabs provided by Animal Genetics. Ensure that the dog has not eaten within a few hours of sample collection. Any food particles can inhibit the test. Rub each of the swabs along the inside of the dog's mouth for 10-15 seconds, and allow the swabs to dry thoroughly. Label the provided envelope with the dog's name, and place the swab inside it. Download and complete a submission form for each sample and send along with payment to Animal Genetics for testing.
Results
Results are given using the following symbolic notation:
KB-Allele Results:
KB/KB | The dog carries two copies of the dominant "KB" allele. The dog will be not be brindled or express his agouti phenotype. The dog will always pass on a copy of the "KB" allele to all offspring. |
KB/n | One copy of the dominant black allele was detected. The dog will be dominant black, and will not express his agouti phenotype. The dog could pass on this allele, or either the brindle or fawn allele, to any offspring. |
n/n | The dog does not carry the dominant black mutation. The dog's coat color will be determined by the agouti gene, and may be brindled or not brindled. |
Want to know the difference between a RED Pomeranian and an Orange Pomeranian? Canine Color Testing- E Locus | |||||||||||||||||||||||||
Description MC1R, also known as the extension gene, controls production of pigment in melanocytes. The dominant form of the gene, the "E" allele, allows the dog to produce eumelanin, which is a black pigment. A mutation in the MC1R gene causes the pigment-producing cells to only produce phaeomelanin, which is a yellow pigment. This form of the gene is represented as the "e" allele. The "e" allele is recessive, meaning that a dog must have two copies of the MC1R mutation to express the solid yellow coat color. A third allele exists in the extension gene, "EM" which is also dominant. This causes the dog to have a black mask on their face, also known as a melanistic mask. This allele acts similarly to the "E" allele, in that it causes a black-based coat. Because it is dominant, a dog only needs one copy of the "EM" allele to express this trait. In solid black dogs with a copy of the "EM" allele, the mask is hidden, however, it can still pass on the melanistic mask to future offspring. The "ee" genotype can vary in expression between different breeds. In some breeds, the difference between a black or brown dog and a yellow dog is obvious, such as in Labrador Retrievers. However, in other breeds, such as Cocker Spaniels, this difference may be more subtle. Other breeds such as pomeranians express the "ee" phenotype as a red color. It is important to note that the Extension gene is only one of 4 important genes in determining the coat color of a canine. The dogs color can vary greatly with different alleles at other gene. Dogs that are "ee" will always be yellow (base), however, there is a great deal of variation of dogs that are "EE" or "Ee", depending on the B-Locus, A-Locus, and K-Locus.
Cost
E-Allele Results:
EM-Allele Results:
Combination Test Results:
|
HORSE COLOR GENETICS
Homozygous creams ("double dilutes")


When a horse is homozygous, meaning it has two copies of the cream allele, the strongest color dilution occurs.
- Cremellos are homozygous cream chestnuts, and have a cream colored body with a cream or white mane and tail.
- Perlinos are homozygous cream bays, which also have a cream-colored body but a mane and tail that may be somewhat more reddish in color than a cremello.
- Smoky Creams are homozygous cream blacks, and very difficult to visually distinguish from cremellos or perlinos.
All three shades can be difficult to distinguish from one another, and are oftentimes only firmly identified after a DNA test. While both red and black pigments are turned cream, the black pigment retains a little more color and tends to have a reddish or rusty tint.[7] Thus all-red coats are turned all-ivory, all-black coats are turned all-rusty cream, and bay coats have ivory bodies with slightly darker points.[3]

{kind=link}
{kind=link}
Horses with two copies of the cream allele can be collectively called double-dilutes, homozygous creams, or blue-eyed creams, and they share a number of characteristics. The eyes are pale blue, paler than the unpigmented blue eyes associated with white color or white markings, and the skin is rosy-pink. The true, unpigmented pink skin associated with white markings is clearly visible against the rosy-pink skin of a double-dilute, especially when their coat is wetted down. The palest shades of double-dilute coats are just off-white, while the darkest are distinctly rust-tinged. Their coats may be described as nearly white[1] or ivory[3] in addition to cream.
The off-white coat, pale blue eyes, and rosy pink skin distinguish the coats of double-dilutes from those of true white horses. True white horses have unpigmented skin and hair due to the incomplete migration ofmelanocytes from the neural crest during development.[citation needed]
No health defects are associated with the cream gene. This is also true of the normal variations in skin, hair and eye color encoded on the human MATP gene.[8]True white coat coloring can be produced by at least half a dozen known genes, and some are associated with health defects. Some genes which encode a white or near-white coat when heterozygous, popularly called "dominant white," may be lethal in homozygote embryos.[9] Another specific mutation on the endothelin receptor type B (EDNRB) gene is associated with theframe overo pattern produces Lethal white syndrome if homozygous, but carriers can be identified with a DNA test.